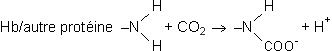|
|
|
|
Présentation généraleLe sang exerce une fonction respiratoire chez la plupart des animaux, même chez de nombreuses espèces ne disposant pas de système respiratoire à proprement parler. Le sang sert en effet très souvent de transporteur d'O2 et de CO2, que ces gaz diffusent à travers un épithélium respiratoire spécialisé ou non. Dans la plupart des cas, l'oxygène n'est pas que dissout dans le sang ; il est fixé sur des molécules particulières servant de transporteurs, augmentant ainsi largement la quantité d'O2 pouvant se trouver dans un volume donné de sang (pouvoir oxyphorique). Ce n'est pas le cas pour le CO2 qui lui est transporté dans le sang largement sous forme de HCO-3.
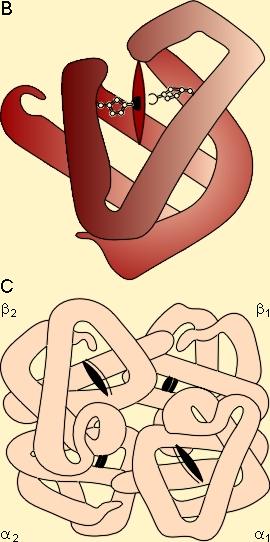
Le transport de l'oxygèneL'oxygène est prélevé dans le milieu ambiant (air) par les poumons lors de la respiration. L'oxygène circule peu dans le sang sous forme gazeuse mais principalement sous forme d'un composé chimique appelé hémoglobine (Hb). La part de l'oxygène transporté sous forme dissoute dans le plasma n'excède pas 0,2 ml pour 100 ml de sang alors que celle transportée par l'Hb atteint 20 ml pour 100 ml de sang, c'est-à-dire 100 fois plus ! Constituant essentiel de l'hématie (globule rouge), cette protéine possède la capacité de fixer l'oxygène destinée à la respiration cellulaire, d'une manière réversible, et d'en assurer le transport du milieu extérieur aux cellules de l'organisme. Cette structure chimique complexe - qui donne la couleur rouge au sang - a pour base 4 atomes de fer auxquels peuvent s'accrocher indifféremment des molécules d'oxygène ou de gaz carbonique. L'hémoglobine fournit 33% du poids des globules rouges. Chez l'homme, le taux normal d'hémoglobine est de 14 à 16 g/dl de sang. La quantité d'hémoglobine présente dans la totalité du sang circulant d'un homme de 70 kg est estimée à 750 g.L'hémoglobine est une hétéroprotéine constituée de globine, et d'une petite molécule non protéique, l'hème :
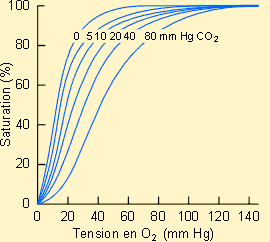
Pouvoir oxyphorique de l'hémoglobineLa charge en oxygène du sang va dépendre de la quantité de molécules
transporteuses qu'il contient et aussi de l'affinité de ces molécules pour
l'oxygène ainsi que de la concentration (pression partielle) de l'O2
dans le sang. On remarque que la cinétique est d'allure sigmoïde, indiquant une
modification d'affinité apparente de l'hémoglobine pour l'O2 en
fonction de sa disponibilité dans le milieu. Cet effet est en fait dû à la
prise en charge de l'O2 sur les sous-unités constitutives de
l'hémoglobine suivant un mode coopératif. Notons par ailleurs que la pression
partielle en O2 du sang artériel est de 95 mmHg alors que celle du
sang veineux n'est que de 40 mmHg. La charge du sang en oxygène sera donc plus
importante dans le sang artériel que dans le sang veineux ou, en d'autres
termes, le sang artériel arrivant aux tissus pourra relarguer une certaine
quantité d'oxygène. Il est intéressant de noter ici que les changements
d'affinité en fonction de la teneur en O2 du milieu vont favoriser
le relarguage de l'O2 aux tissus en conditions physiologiques. Il est
clair dans ce contexte que la plus ou moins bonne capacité à délivrer
l'oxygène aux tissus ou à fixer l'oxygène au niveau des échangeurs
respiratoires dépendra de l'affinité du transporteur pour l'O2 Effet BohrLes variations de pH sanguin ou de pression partielle en CO2 provoquent des déplacements des courbes de dissociation de l'O2 dans le sang. Cet effet a été mis en évidence pour la première fois en 1904 par Bohr, Hasselbalch et Krogh, d'où le nom d'effet Bohr qu'il porte encore aujourd'hui. Comme le montre les figures suivantes, une augmentation de PCO2 ou une diminution de pH provoque un déplacement plus ou moins marqué de la courbe vers la droite. Il y a donc un meilleur relarguage de l'O2 au niveau tissulaire. Ce type d'effet est particulièrement intéressant à considérer dans le cadre d'une activité musculaire importante, condition amenant une augmentation notable de production de CO2 parfois accompagnée d'une production d'acide lactique et dans laquelle la demande en oxygène augmente largement.
Transport du gaz carboniqueContrairement à ce qui se passe pour l'oxygène, il n'existe pas de transporteur fixant de façon spécifique le CO2 dans le sang. Le CO2 va donc se retrouver, pour une petite part, sous forme libre, dissout dans le plasma et le liquide intracellulaire des éléments figurés sanguin. Cette fraction est loin d'être négligeable puisque ces éléments, avec parmi eux essentiellement les globules rouges, peuvent représenter quelque 40 % du volume sanguin total. Une autre petite partie du CO2 sera fixée aux protéines du plasma ou des globules rouges, essentiellement l'hémoglobine dans ce dernier cas. Il s'agit ici d'une liaison carbamino-protéique faisant intervenir les groupements N terminaux :
L'essentiel du CO2 (90 % chez l'homme) se trouve en fait sous forme de bicarbonate dans le plasma et les globules rouges.
Distribution du CO2 sanguin chez l'homme, en % du total
Un point remarquable concernant la liaison du CO2 à l'hémoglobine est le fait que l'hémoglobine désoxygénée peut fixer plus de CO2 que l'hémoglobine oxygénée. Ce phénomène, connu sous le nom d'effet Haldane présente un certain intérêt physiologique puisque l'hémoglobine ayant déchargé une partie de son oxygène au niveau des tissus pourra fixer plus de CO2. Ce CO2 sera par contre libéré plus facilement au niveau des échangeurs respiratoires, l'hémoglobine se chargeant en oxygène.
| |||||||||||||||||||||||||
|
(C) 2000 ObNet Les informations de ce site peuvent être copiées dans un but d'information non commerciale après accord à demander au Webmaster. Les photographies sont la propriété de leurs auteurs et ne peuvent être dupliquées sans leur autorisation. |


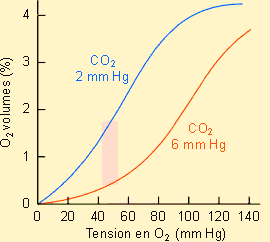
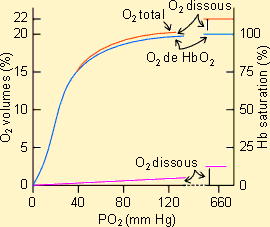 La figure ci-jointe montre, à titre d'exemple, la relation qui
existe entre la concentration en O2 du milieu et la pression
partielle de l'O2 dans le sang chez un mammifère. Comme on le voit,
la quantité d'O2 totale fait intervenir une fraction d'O2
dissout dans le plasma sanguin et une fraction, largement plus importante, d'O2
lié à l'hémoglobine. La fraction O2 dissout augmente
régulièrement avec l'augmentation de la pression artérielle en O2
(loi de Henry) alors de la fraction O2 de l'hémoglobine montre une
cinétique à saturation avec un plateau aux environs de 20 volumes %. La
quantité d'O2 totale prise en charge par le sang en conditions de
saturation définit ce qu'il est convenu d'appeler le pouvoir oxyphorique du
sang.
La figure ci-jointe montre, à titre d'exemple, la relation qui
existe entre la concentration en O2 du milieu et la pression
partielle de l'O2 dans le sang chez un mammifère. Comme on le voit,
la quantité d'O2 totale fait intervenir une fraction d'O2
dissout dans le plasma sanguin et une fraction, largement plus importante, d'O2
lié à l'hémoglobine. La fraction O2 dissout augmente
régulièrement avec l'augmentation de la pression artérielle en O2
(loi de Henry) alors de la fraction O2 de l'hémoglobine montre une
cinétique à saturation avec un plateau aux environs de 20 volumes %. La
quantité d'O2 totale prise en charge par le sang en conditions de
saturation définit ce qu'il est convenu d'appeler le pouvoir oxyphorique du
sang.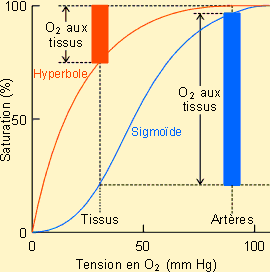 mais
également de la pression partielle en O2 prévalant localement. Elle
sera aussi fonction d'autres paramètres, comme la teneur en CO2 du
sang ou sa température (voir plus loin) mais aussi la vitesse d'écoulement et
les possibilités de diffusion. Dans ce cadre, ce serait une erreur de croire
que le sang d'un organisme délivrera l'entièreté de l'oxygène qu'il contient
aux tissus. Le relarguage au niveau tissulaire tout comme d'ailleurs la charge
au niveau respiratoire seront en effet fonction non seulement de l'affinité du
transporteur pour l'O2 mais encore des surfaces de diffusion et des
gradients en O2 existants ainsi que de la vitesse de perfusion des
organes ; ces paramètres étant eux-mêmes variables en fonction de nombreux
facteurs. Ainsi chez l'homme au repos par exemple, si l'hémoglobine est
saturée à 95-97 % au niveau pulmonaire, elle n'est désaturée que de 25 à 40
% au niveau des tissus. Ces dernières valeurs sont éminemment variables,
dépendant de la demande énergétique de l'organe envisagé ; en moyenne
l'hémoglobine dans le sang veineux de retour reste saturée à près de 70 % ;
ce qui laisse une marge importante permettant de répondre à une forte demande.
mais
également de la pression partielle en O2 prévalant localement. Elle
sera aussi fonction d'autres paramètres, comme la teneur en CO2 du
sang ou sa température (voir plus loin) mais aussi la vitesse d'écoulement et
les possibilités de diffusion. Dans ce cadre, ce serait une erreur de croire
que le sang d'un organisme délivrera l'entièreté de l'oxygène qu'il contient
aux tissus. Le relarguage au niveau tissulaire tout comme d'ailleurs la charge
au niveau respiratoire seront en effet fonction non seulement de l'affinité du
transporteur pour l'O2 mais encore des surfaces de diffusion et des
gradients en O2 existants ainsi que de la vitesse de perfusion des
organes ; ces paramètres étant eux-mêmes variables en fonction de nombreux
facteurs. Ainsi chez l'homme au repos par exemple, si l'hémoglobine est
saturée à 95-97 % au niveau pulmonaire, elle n'est désaturée que de 25 à 40
% au niveau des tissus. Ces dernières valeurs sont éminemment variables,
dépendant de la demande énergétique de l'organe envisagé ; en moyenne
l'hémoglobine dans le sang veineux de retour reste saturée à près de 70 % ;
ce qui laisse une marge importante permettant de répondre à une forte demande.