|
|
|
|
Les musclesLe corps humain compte 639 muscles représentant environ 40% de son poids. Le plus volumineux est le muscle fessier ; le plus petit (à peine 1 millimètre de long) est le muscle stapédien situé à l'intérieur de l'oreille. Leur importance est très inégale : certains peuvent être enlevés sans dommage, alors que d'autres, comme le muscle cardiaque, sont vitaux.Ils sont composés de 75% d'eau, 21% de protéines (myosine), 1% de glycogène (réserve d'énergie), de sels minéraux, de composés azotés et phosphorés jouant un rôle important dans la contraction musculaire. Tous les muscles sont caractérisés par leur capacité à se contracter ; mais ils ne peuvent fonctionner seuls et dépendent étroitement du système nerveux qui les contrôle. Marcher, courir, manger, cligner de l'œil ou «simplement» respirer est bien plus compliqué qu'il n'y paraît !
ContractionLe muscle est constitué d'un groupement de faisceaux, formés eux-mêmes d'un ensemble de fibres musculaires, serrées les unes contre les autres. Chacune de ces fibres est une cellule musculaire de très grande taille, qui comprend plusieurs noyaux. Une fibre musculaire peut atteindre plusieurs dizaines de centimètres dans les grands muscles du dos ou des membres inférieurs. À l'intérieur de la fibre sont alignés des faisceaux de myofibrilles, protéines contractiles qui donnent au muscle le pouvoir de se contracter, puis de se décontracter. Les myofibrilles sont constituées de filaments, parmi lesquels on distingue deux types : les filaments fins d'actine et les filaments épais de myosine. Au moment de la contraction, les filaments d'actine et de myosine glissent les uns contre les autres, de façon à raccourcir le muscle. Les molécules de myosine, plus épaisses, sont équipées de têtes spéciales, qui viennent s'accrocher aux molécules d'actine. Le mouvement musculaire entraîne un certain nombre de phénomènes, qui sont encore loin d'être tous expliqués.
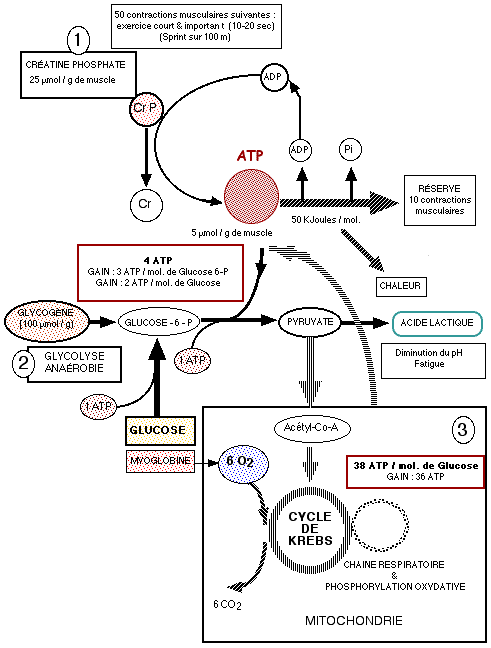
Les carburants du muscleL'énergie mécanique de la contraction musculaire provient directement de l'énergie chimique stockée dans une molécule biologique : l'ATP. Pendant l'activité musculaire, la régénération de l'ATP se fait suivant 3 voies :
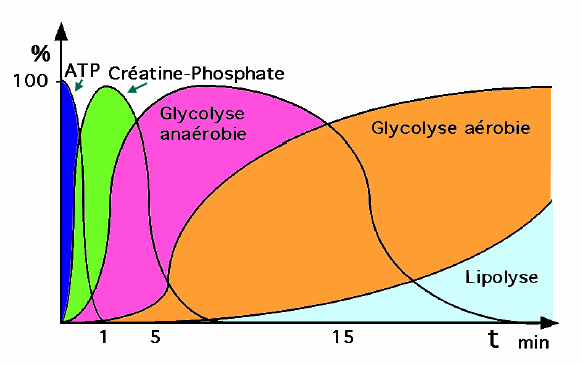
Réserve d'ATPL'organisme trouve sa première source énergétique dans l'ATP présent dans le muscle qui lui donne deux à trois secondes d'énergie. A aucun moment, l'organisme n'emmagasine plus de 85 g d'ATP. Cette énergie ne fait pas appel à l'oxygène, elle est anaérobie et ne laisse aucun déchet. Cette énergie possède le plus fort pouvoir explosif mais est éphémère. Du fait que l'ATP ne peut être fourni par voie sanguine ni par les tissus, il doit être continuellement resynthétisé dans la cellule. Réaction couplée de l'ADP avec la créatine phosphate (1)Dès l'épuisement des réserves d'ATP, un système de production rapide d'ATP se met en place, en attendant que les voies métaboliques s'adaptent à la demande accrue d'ATP. La filière énergétique anaérobique alactique est une source immédiatement disponible. Elle fait appel à l'action de deux enzymes:
L'AMP ou adénosine monophosphate semble par sa concentration favoriser le développement et l'action d'autres enzymes. Une puissance musculaire maximale peut ainsi être maintenue pendant 10 à 15 secondes (sprint sur 100 m). Respiration cellulaire anaérobie (2)Pour des exercices musculaires de plus longue durée, le glycogène musculaire doit être dégradé. Les réserves de glycogène du muscle sont transformées en acide lactique via le glucose-6-phosphate, avec production de 3 molécules d'ATP par molécule de sucre (faible rendement énergétique) au cours de la glycogènolyse. La glycolyse anaérobie commence plus tardivement que la dégradation de la créatine phosphate (au maximum après 30 secondes) et produit de l'ATP 2,5 fois plus vite que la voie aérobie. Ainsi, lorsqu'il faut de grandes quantités d'ATP pendant de courtes périodes d'activité musculaire soutenue (30-40 secondes), la voie anaérobie en fournit une grande partie. Ensemble, les réserves d'ATP et de créatine phosphate et le système glycogénolyse-acide lactique peuvent entretenir une activité musculaire pendant presque une minute. Respiration cellulaire aérobie : la phosphorylation oxydative du glucose et des acides gras (3)Pour que l'effort soit maintenu, il faut absolument qu'il y ait hydrolyse aérobie du glucose et des acides gras. Lors des contractions lentes ou au repos, la plus grande partie de l'approvisionnement en ATP est assurée par la respiration cellulaire aérobie, qui utilise l'énergie fournie par la dégradation des acides gras. Lorsque les muscles se contractent de façon plus soutenue, c'est le glucose qui devient la principale source d'énergie. La respiration cellulaire aérobie se déroule dans les mitochondries; elle nécessite la présence d'oxygène et fait intervenir une suite de réactions complexes (cycle de Krebs - chaîne respiratoire de transport d'électrons - phosphorylation oxydative). Dans la chaîne respiratoire, les atomes d'hydrogène enlevés au cours de la dégradation des combustibles finissent par être combinés avec l'oxygène moléculaire, et l'énergie libérée est utilisée pour lier les groupements phosphate inorganique (Pi) à l'ADP. Globalement, l'oxydation complète d'une molécule de glucose en CO2 et en eau fournit 36-38 molécules d'ATP (rendement énergétique élevé).
Propriétés physiologiques des musclesExcitabilitéLe muscle réagit à des excitants mécaniques (piqûre, choc, pincement, blessure), thermiques (variations brusques de température), chimiques (acides et alcalis faibles, chlorure de sodium, glycérine), électriques (variations brusques d'intensité d'un courant continu, d'un courant induit, décharges d'un condensateur), physiologiques (influx nerveux amené par les nerfs). ÉlasticitéCelle du muscle proprement dit est assez faible. Elle est due à l'allongement des disques clairs du muscle.
Mais les muscles sont la plupart du temps en relation avec des éléments tendineux, conjonctifs ou chitineux dont l'élasticité permet d'emmagasiner une partie, parfois très importante, de l'énergie développée par un muscle dans le cadre d'un mouvement pour la restituer dans le cadre d'un autre mouvement. On se rend facilement compte de l'intérêt des systèmes élastiques en effectuant par exemple des flexions répétées des genoux. Il est en effet nettement plus facile de se redresser si on n'exécute pas de pose en position accroupie. On met en effet ainsi à profit un rebond élastique dû essentiellement à la tension développée par les muscles extenseurs des jambes pour contrôler la descente et emmagasinée dans les tendons de ces muscles. La pose aboutit au relâchement de ces muscles et donc, à la perte de cette tension et du rebond élastique. On a ainsi pu mettre en évidence chez l'homme effectuant vingt flexions des genoux avec ou sans pose, une diminution de 20 % de la consommation d'oxygène et une augmentation de l'efficacité du mouvement de près de 40 % ContractilitéOn distingue les formes de contraction suivantes :
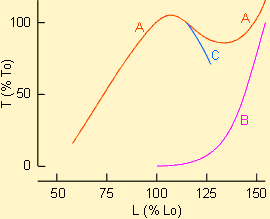
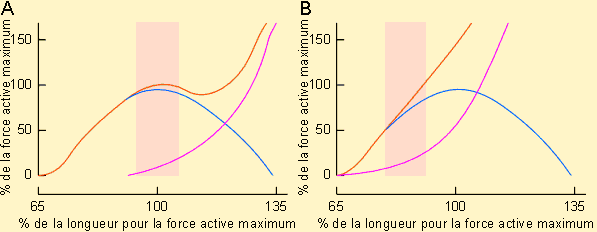
En étudiant un muscle strié en mode de contraction isométrique, il est possible de déterminer facilement la tension qu'il développe à partir de différentes longueurs de départ. On s'aperçoit que, dans ces conditions, la tension maximum est obtenue pour une longueur égale ou très légèrement supérieure à la longueur de repos. En deçà de cette longueur, la tension développée diminue; au-delà, la tension peut diminuer puis augmenter jusqu'à un certain point, variable en fonction du muscle considéré.
Ces évolutions se comprennent aisément en considérant l'organisation des fibres d'actine et de myosine dans la mécanique contractile d'une part et la présence d'éléments élastique dans le muscle d'autre part. L'étirement du muscle va en effet étirer les éléments élastiques qui en font partie (tendons, matrices extracellulaires, protéines particulières de la masse myofibrillaire telle la titine...) ce qui va rendre compte du développement d'une certaine tension dite passive n'ayant rien à voir avec la tension active due à l'activation de la mécanique contractile et au raccourcissement des sarcomères. Cette tension sera d'autant plus importante que les fibres musculaires sont étirées. L'importance de la tension active sera fonction du chevauchement plus ou moins important des filaments d'actine et de myosine et donc de leurs possibilités d'interaction. Il est intéressant de remarquer dans ce cadre que les membranes Z des myofibrilles de certains muscles (fibres géantes de barnacle par exemple) présentent des trous qui permettent à la myosine de pénétrer dans le sarcomère voisin; ce qui paraît procurer une plage de travail plus large qu'un muscle normal où la myosine vient buter sur les limites des sarcomères. Le muscle cardiaque est moins extensible que le muscle squelettique. Pour un même allongement, la tension passive qu'il développe sera donc plus grande. Les courbes tension - longueur ne montrent donc pas de plateau. De plus, le muscle cardiaque travaille dans la partie ascendante de la tension active alors que le muscle squelettique travaille essentiellement dans le plateau.
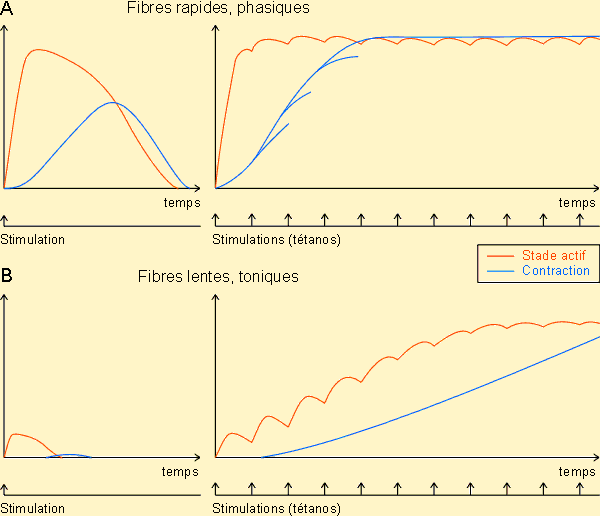
L'intérêt majeur des fibres lisses réside dans le fait qu'elles peuvent subir des étirements très importants tout en conservant leurs possibilités de contraction. Ce n'est pas le cas d'un muscle strié où, comme nous l'avons vu, un étirement important rend la contraction impossible. Cette caractéristique rend le muscle lisse particulièrement intéressant au niveau de structures telles que la vessie, l'estomac ou l'intestin qui peuvent être soumises à des distensions importantes. La tétanisation est un état de contraction permanente du muscle ; il est imparfait lorsque, à la suite d'excitations nombreuses et rapprochées, le muscle est sollicité à nouveau avant de s'être complètement relâché. Le courant alternatif de basse fréquence provoque une tétanisation. La rhéobase (du grec rhéos , courant) : seuil minimal d'intensité du courant nécessaire pour qu'un muscle puisse se contracter ; varie selon chaque muscle. La chronaxie est le temps minimal pendant lequel il faut faire passer un courant d'intensité double de la rhéobase pour obtenir une contraction du muscle. ÉnergieVient de réactions chimiques. Le muscle au travail consomme de l'oxygène (jusqu'à 30 fois plus qu'au repos) et du glycogène (accumulé dans le muscle au repos à partir du glucose du sang) pour fournir de l'énergie, de l'eau, du gaz carbonique (dans les mêmes proportions que l'oxygène consommé) et de la chaleur. Chaleur d'activation, types de muscle et propriétés contractilesComme tout processus physiologique, la contraction musculaire s'accompagne d'un dégagement de chaleur qui peut être facilement mesuré si le muscle est suffisamment important. La première manifestation de ce dégagement de chaleur suit directement la stimulation du muscle. Elle atteint son maximum à un moment où soit le mouvement, soit la tension est à peine décelable; elle constitue donc un signe avant-coureur de la contraction proprement dite, probablement à mettre en rapport avec l'initiation des interactions actine - myosine, d'où son nom de chaleur d'activation. Elle serait dans ce contexte significative de la mise du système au stade actif. En suivant cette production de chaleur et en la mettant en rapport avec l'évolution de l'activité musculaire, on met en évidence deux types de fibres musculaires très différentes :
Dans un cas, les fibres répondent à un stimulus par le développement rapide d'un stade actif important suivi d'une contraction elle-même importante. Dans l'autre cas, les fibres ne présentent qu'un stade actif peu important et une contraction à peine significative. Soumettons maintenant ces deux types de fibres à des stimulations répétées. Dans le premier cas, on obtient très vite par sommation un stade actif et une contraction maximum, appelée également tétanos. Dans le deuxième cas, la contraction et le stade actif n'évoluent que lentement vers un maximum.
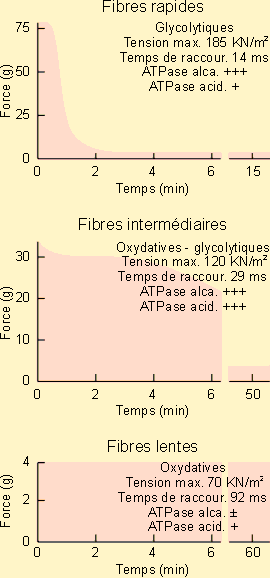
Les premières fibres ont été appelées fibres rapides, phasiques ou de type 2. Elles correspondent à des muscles de coloration en général assez pâles d'où leur nom de muscles blancs. Les secondes ont été appelées fibres lentes, tonique ou de type 1. Elles sont les plus abondantes dans des muscles de coloration foncée, appelés muscles rouges. La coloration est ici essentiellement due à la présence de myoglobine en quantité importante à l'intérieur des fibres. On y trouve également beaucoup plus de mitochondries que dans les fibres rapides mais moins de myofibrilles. Leur vascularisation est également plus importante de même que leurs réserves lipidiques. Elles sont en fait organisées pour développer une puissance assez faible pendant longtemps, en fonctionnant de manière aérobie sur leurs réserves lipidiques. Elles montrent dans ce cadre une grande résistance à la fatigue. Leur myosine paraît porter des ATPases à action lente en petite quantité; ce qui serait responsable de l'activation et de la contraction lente observée à leur niveau. Les fibres rapides possèdent par contre en grandes quantités des ATPases à action rapide rendant compte de leur vitesse de réaction. Elles n'ont que peu de mitochondries et de myoglobine mais plus de myofibrilles contractiles. Elles sont également moins richement vascularisées que les fibres lentes. Elles n'ont que peu de réserves lipidiques mais présentent par contre des quantités appréciables de glycogène. Elles sont donc organisées pour fonctionner puissamment mais brièvement en puisant leur énergie dans une séquence anaérobie. Ces fibres montreront rapidement des signes de fatigue et d'accumulation d'acide lactique. Des études physiologiques et histologiques plus fines ont mis en évidence un troisième type de fibres, dites fibres intermédiaires. Elles ont des ATPases des deux types avec moins de myofibrilles mais plus de mitochondries et de myoglobine que les fibres rapides. Elles pourront donc développer une force moins grande que celles-ci mais seront plus résistantes à la fatigue. On a ainsi mis en évidence différentes sortes de fibres plus ou moins rapides et plus ou moins fatigables chez tous les animaux. Un muscle ne comprend que très rarement un seul type de fibres. Il est en général constitué de l'association de 2 ou des 3 types de fibres, le type de fibres prédominant donnant les caractéristiques essentielles du muscle. La plupart des muscles sont à prédominance de fibres intermédiaires et lentes, conçus donc pour pouvoir maintenir des activités soutenues pendant des périodes assez longues. Les muscles essentiellement rapides (blancs ou roses) sont rares.
Activité physique et dépense énergétiqueVoici une idée de l'énergie dépensée au cours de différentes activités sportives (en Kcal/h) :
Ces valeurs sont très approximatives et peuvent varier dans un rapport de 1 à 4 suivant la tension nerveuse, l'entraînement, l'intensité, etc...
|
|
(C) 2000 ObNet Les informations de ce site peuvent être copiées dans un but d'information non commerciale après accord à demander au Webmaster. Les photographies sont la propriété de leurs auteurs et ne peuvent être dupliquées sans leur autorisation. |